



Sterol Disorders Panel
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Panel CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 13319 | Genes x (12) | 81479 | 81405(x1), 81479(x23) | $990 | Order Options and Pricing |
Pricing Comments
We are happy to accommodate requests for testing single genes in this panel or a subset of these genes. The price will remain the list price. If desired, free reflex testing to remaining genes on panel is available. Alternatively, a single gene or subset of genes can also be ordered via our Custom Panel tool.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
In humans, de novo synthesis of cholesterol occurs via the isoprenoid/cholesterol synthesis pathway. The isoprenoid compounds generated via this pathway play important roles in processes such as cell growth and differentiation, protein glycosylation, and signal transduction (Waterham and Clayton. 2016). In most eukaryotic cells, cholesterol, which is the final product in this biochemical pathway, is a key component of cell membranes and lipid rafts, is important in myelin formation in the brain, spinal cord, and peripheral nervous system, and is the immediate precursor of steroid hormones, vitamin D, and bile acids. In addition, cholesterol interacts with proteins that are involved in control of embryonic development (He et al. 2011. PubMed ID: 21285510; Jira. 2013. PubMed ID: 23622407). Consequently, defects in enzymes in the isoprenoid/cholesterol synthetic pathway can lead to abnormalities in morphogenesis, growth delay, and developmental delays (Waterham and Clayton. 2016; Herman. 2003. PubMed ID: 12668600; Jira. 2013. PubMed ID: 23622407).
Disorders of isoprenoid/cholesterol synthesis are clinically heterogeneous. Phenotypes can range in severity from very mildly affected to severe disorders that are lethal in utero (Waterham and Clayton. 2016; Herman. 2003. PubMed ID: 12668600; Jira. 2013. PubMed ID: 23622407). Clinical presentations of disorders of isoprenoid/cholesterol synthesis may include but are not limited to: dysmorphic facial features, cataracts, mild to severe skin abnormalities (such as ichthyosiform erythroderma), mild to severe skeletal abnormalities (such as severe limb defects or skeletal dysplasias, kyphosis, and/or scoliosis), genital abnormalities (such as hypospadias or ambiguous genitalia), digit abnormalities (2-3 syndactyly of the toes is particularly common, although other abnormalities may occur), growth delay and/or failure to thrive, intellectual and/or developmental delay, behavioral abnormalities, and autism spectrum disorders. Patients often present prenatally or in infancy or early childhood (Waterham and Clayton. 2016; Wassif et al. 1998. PubMed ID: 9634533; Herman. 2003. PubMed ID: 12668600; Sikora et al. 2006. PubMed ID: 16761297; He et al. 2011. PubMed ID: 21285510; Jira. 2013. PubMed ID: 23622407; Eroglu et al. 2017. PubMed ID: 28349652; Nowaczyk and Wassif. 2020. PubMed ID: 20301322).
Biochemically, affected patients are often found to have low to normal cholesterol levels and other abnormalities in sterol profiles as measured in plasma, tissues, or cultured patient cells (Waterham and Clayton. 2016; Herman. 2003. PubMed ID: 12668600; He et al. 2011. PubMed ID: 21285510; Jira. 2013. PubMed ID: 23622407; Nowaczyk and Wassif. 2020. PubMed ID: 20301322). Individual disorders are typically associated with a particular abnormality in the sterol profile (for example, elevation of 7-dehydrocholesterol is diagnostic for Smith-Lemli-Opitz syndrome) (Waterham and Clayton. 2016; Porter and Herman. 2011. PubMed ID: 20929975; Coman et al. 2020. PubMed ID: 32027475; Nowaczyk and Wassif. 2020. PubMed ID: 20301322).
Also included in this test are genes associated with cytochrome P450 oxidoreductase (POR) deficiency and the sitosterolemias. Patients with POR deficiency may present similarly to those with a disorder of isoprenoid or cholesterol synthesis, with the most severely affected patients presenting with craniofacial anomalies, skeletal defects, and ambiguous genitalia (Porter and Herman. 2011. PubMed ID: 20929975; Idkowiak et al. 2017. PubMed ID: 20301592). In contrast, the clinical features of sitosterolemia patients are distinct and include hypercholesterolemia (especially in children) with an unexpected lowering of plasma cholesterol level in response to low-fat diet or bile acid sequestrant therapy, tendon or tuberous xanthomas, premature atherosclerosis which can lead to cardiac issues such as aortic valve involvement, myocardial infarction, and sudden death. Patients may also have hematologic abnormalities including hemolytic anemia, abnormally shaped erythrocytes, and/or large platelets, and these may occasionally be the only finding (Myrie et al. 2020. PubMed ID: 23556150). Biochemical testing of patients with POR deficiency or sitosterolemia may show an abnormal sterol profile (Porter and Herman. 2011. PubMed ID: 20929975; Myrie et al. 2020. PubMed ID: 23556150).
Two of the genes in this panel (EBP, associated with X-linked chondrodysplasia punctata 2, CDPX2, and NSDHL, associated with CHILD and CK syndromes) are inherited in an X-linked manner. The majority of affected individuals are female as severe pathogenic variants in these genes are lethal in the hemizygous state. However, some affected males have been reported with variants in each gene, typically in the setting of an aberrant karyotype (such as 47,XXY), somatic mosaicism, or a hypomorphic, mild variant (Waterham and Clayton. 2016). Phenotypes in affected females may vary based on X-inactivation (du Souich et al. 2018. PubMed ID: 21290788; Kumble and Savarirayan. 2020. PubMed ID: 21634086).
The most common disorder covered by this panel is Smith-Lemli-Opitz syndrome (SLOS), with an estimated prevalence of ~1:40,000 in North America (Battaile et al. 2001. PubMed ID: 11161831; Nowaczyk and Wassif. 2020. PubMed ID: 20301322). The other disorders in this panel are much rarer, generally ranging from only a few patients to a few hundred patients reported worldwide.
At the present time, there are limited treatment options available to individuals with these disorders, and management is primarily supportive. Molecular diagnosis may help assess recurrence risks. Dietary supplementation or modification (such as cholesterol supplementation) may have some benefit in certain cases (Myrie et al. 2020. PubMed ID: 23556150; Nowaczyk and Wassif. 2020. PubMed ID: 20301322).
Genetics
Pathogenic variants in twelve genes are associated with the disorders of isoprenoid/cholesterol synthesis, POR deficiency, and the sitosterolemias. All have autosomal recessive inheritance with the exception of the EBP and NSDHL genes, which have X-linked inheritance. To our knowledge, EBP and NSDHL are the only genes in this panel in which de novo variants are commonly reported to cause disease. Pathogenic defects in most of the genes in this panel include missense, nonsense, splicing site variants, small deletions, small insertions/duplications, small indels, and exon-level large deletions, although in a few cases only missense variants have been reported (DHCR24, MSMO1) (Human Gene Mutation Database).
Pathogenic variants in DHCR7 are associated with Smith-Lemli-Opitz syndrome (SLOS), which is the most commonly occurring disorder covered by this panel. DHCR7 encodes 7-dehydrocholesterol reductase, which catalyzes the predominant final step of cholesterol biosynthesis (Wassif et al. 1998. PubMed ID: 9634533; Herman. 2003. PubMed ID: 12668600; Sikora et al. 2006. PubMed ID: 16761297; Jira. 2013. PubMed ID: 23622407; Eroglu et al. 2017. PubMed ID: 28349652). The c.964-1G>C, c.452G>A, and c.1210C>T variants are the most common pathogenic variants associated with a severe phenotype (Battaile et al. 2001. PubMed ID: 11161831; Nowaczyk and Wassif. 2020. PubMed ID: 20301322).
Pathogenic variants in MVK are associated with mevalonic aciduria (MKD-MA) and hyper-IgD and periodic fever syndrome (MKD-HIDS). The MVK gene encodes the enzyme mevalonate kinase, which is involved in isoprenoid synthesis (Waterham and Clayton. 2016).
Pathogenic variants in FDFT1 are associated with squalene synthase deficiency. The FDFT1 gene encodes the enzyme squalene synthase (also called farnesyl-pyrophosphate farnesyl-transferase 1), which catalyzes the conversion of farnesyl-pyrophosphate to pre-squalene diphosphate (Coman et al. 1993. PubMed ID: 32027475). Of note, a deletion within the 5’ UTR of FDFT1 that has been reported to result in decreased promoter activity (Coman et al. 1993. PubMed ID: 32027475).
Pathogenic variants in DHCR24 are associated with desmosterolosis. The DHCR24 gene encodes the enzyme sterol D24-reductase, which catalyzes reduction of the D24 double bond of sterol intermediates (Waterham and Clayton. 2016).
Pathogenic variants in MSMO1 are associated with sterol-C4-methyloxidase (SC4MOL) deficiency. The MSMO1 gene encodes the enzyme sterol-C4-methyl oxidase, which catalyzes demethylation of C4-methylsterols (He et al. 2011. PubMed ID: 21285510). To date, only missense variants have been reported to be causative for SC4MOL deficiency (Human Gene Mutation Database).
Pathogenic variants in LBR are associated with Greenberg skeletal dysplasia (also known as hydrops-ectopic calcification-moth-eaten (HEM) skeletal dysplasia). The LBR gene encodes the lamin B receptor, which in the cholesterol synthesis pathway catalyzes the reduction of D14 double bonds in early sterol intermediates (Waterham and Clayton. 2016). To date, missense, small deletion, and small indel variants have been reported to be causative for Greenberg skeletal dysplasia (Human Gene Mutation Database). Of note, heterozygous pathogenic variants in LBR are also associated with autosomal dominant Pelger-Huet anomaly, which is a disorder of leukocyte development. Individuals homozygous for Pelger-Huet associated variants may present with minor skeletal abnormalities and developmental delay (Waterham and Clayton. 2016).
Pathogenic variants in SC5D are associated with lathosterolosis. The SC5D gene encodes the 3b-hydroxysterol D5-desaturase enzyme, which introduces a C5-C6 double bond in lathosterol to produce 7-dehydrocholesterol (Waterham and Clayton. 2016).
The POR gene encodes the cytochrome P450 oxidoreductase (POR), which acts as an electron donor to many cytoplasmic P450 enzymes, including some involved in cholesterol or steroid hormone synthesis (Porter and Herman. 2011. PubMed ID: 20929975). The p.Ala287Pro and p.Arg457His variants account for ~40% and 50% of pathogenic alleles among European and Japanese patients, respectively (Porter and Herman. 2011. PubMed ID: 20929975).
Sitosterolemia is caused by pathogenic variants in ABCG5 or ABCG8, which encode the proteins sterolin-1 and sterolin-2. Together, these proteins form a heterodimeric transporter involved in absorption of dietary cholesterol and sitosterol, as well as excretion of cholesterol and sitosterol into the bile (Myrie et al. 2020. PubMed ID: 23556150).
The EBP and gene is associated with X-linked chondrodysplasia punctata 2 (CDPX2, also called Conradi-Hünermann syndrome). The EBP gene encodes the enzyme 3-b-hydroxysteroid-D8-7-isomerase, which converts 8(9)-cholestenol to lathosterol during cholesterol biosynthesis (Kumble and Savarirayan. 2020. PubMed ID: 21634086). Pathogenic variants may be either inherited or occur de novo (Kumble and Savarirayan. 2020. PubMed ID: 21634086).
The NSDHL gene is associated with the X-linked disorders CHILD syndrome (congenital hemidysplasia with ichthyosiform erythroderma and limb defects) which occurs nearly exclusively in females, and CK syndrome, which has been reported in several males (du Souich et al. 2018. PubMed ID: 21290788). Pathogenic variants may be either inherited or occur de novo (du Souich et al. 2018. PubMed ID: 21290788).
For the majority of genes on this panel, large copy number variants (gross deletions or duplications/insertions) are a rare cause of disease. An exception to this is NSDHL, in which ~11% of reported variants have been multi-exon or full-gene deletion (du Souich et al. 2018. PubMed ID: 21290788).
See individual gene summaries for more information about molecular biology of gene products and spectra of pathogenic variants.
Clinical Sensitivity - Sequencing with CNV PGxome
As no large cohort has been described to date that has been tested for this subset of genes, clinical sensitivity is difficult to estimate. However, clinical sensitivity can be estimated for many of the individual genes. Pathogenic variants in DHCR7 were identified in >96% of SLOS patients (Nowaczyk and Wassif. 2020. PubMed ID: 20301322). Pathogenic variants in MVK were identified in 88-100% of patients with biochemical evidence of impaired mevalonate kinase activity and suspected MA or HIDS (Mandey et al. 2006. PubMed ID: 16835861; Bader-Meunier et al. 2010. PubMed ID: 21708801). Pathogenic variants in POR were identified in 94.5% of patients with cytochrome P450 oxidase deficiency (Idkowiak et al. 2017. PubMed ID: 20301592). Pathogenic variants in either ABCG5 or ABCG8 were identified in >95% of patients with sitosterolemia (Myrie et al. 2020. PubMed ID: 23556150). Pathogenic variants have been reported in 100% of patients with NSDHL-related disorders (du Souich et al. 2018. PubMed ID: 21290788) and CDPX2 (Kumble and Savarirayan. 2020. PubMed ID: 21634086).
For desmosterolosis, squalene synthase deficiency, Greenberg skeletal dysplasia, SC4MOL deficiency, and lathosterolosis, clinical sensitivity cannot be estimated because only a small number of patients with DHCR24, FDFT1, LBR, MSMO1, or SC5D pathogenic variants have been reported.
Large deletions or duplications have been reported in ABCG5, ABCG8, DHCR7, MVK, NSDHL, and POR (Human Gene Mutation Database).
Testing Strategy
This test is performed using Next-Gen sequencing with additional Sanger sequencing as necessary.
This panel provides 100% coverage of all coding exons of the genes plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define coverage as ≥20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Patients who present with low to normal cholesterol and an abnormal sterol profile, measured from plasma, tissue, or cultured patient cells, and/or a clinical presentation that is suggestive of a sterol disorder would be good candidates for this test.
Patients who present with low to normal cholesterol and an abnormal sterol profile, measured from plasma, tissue, or cultured patient cells, and/or a clinical presentation that is suggestive of a sterol disorder would be good candidates for this test.
Genes
| Official Gene Symbol | OMIM ID |
|---|---|
| ABCG5 | 605459 |
| ABCG8 | 605460 |
| DHCR24 | 606418 |
| DHCR7 | 602858 |
| EBP | 300205 |
| FDFT1 | 184420 |
| LBR | 600024 |
| MSMO1 | 607545 |
| MVK | 251170 |
| NSDHL | 300275 |
| POR | 124015 |
| SC5D | 602286 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
Related Test
| Name |
|---|
| PGxome® |
Citations
- Bader-Meunier et al. 2011. PubMed ID: 21708801
- Battaile et al. 2001. PubMed ID: 11161831
- Coman et al. 2020. PubMed ID: 32027475
- du Souich et al. 2011. PubMed ID: 21290788
- Eroglu et al. 2017. PubMed ID: 28349652
- He et al. 2011. PubMed ID: 21285510
- Herman. 2003. PubMed ID: 12668600
- Human Gene Mutation Database (Biobase).
- Idkowiak et al. 2017. PubMed ID: 20301592
- Jira. 2013. PubMed ID: 23622407
- Kumble and Savarirayan. 2020. PubMed ID: 21634086
- Mandey et al. 2006. PubMed ID: 16835861
- Myrie et al. 2020. PubMed ID: 23556150
- Nowaczyk and Wassif. 2020. PubMed ID: 20301322
- Porter and Herman. 2011. PubMed ID: 20929975
- Sikora et al. 2006. PubMed ID: 16761297
- Wassif et al. 1998. PubMed ID: 9634533
- Waterham and Clayton. 2016. Disorders of Isoprenoid/Cholesterol Synthesis. In: Saudubray, Baumgartner and Walter, editors. Berlin: Springer. Inborn Metabolic Diseases: Diagnosis and Treatment, 6th Ed.
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
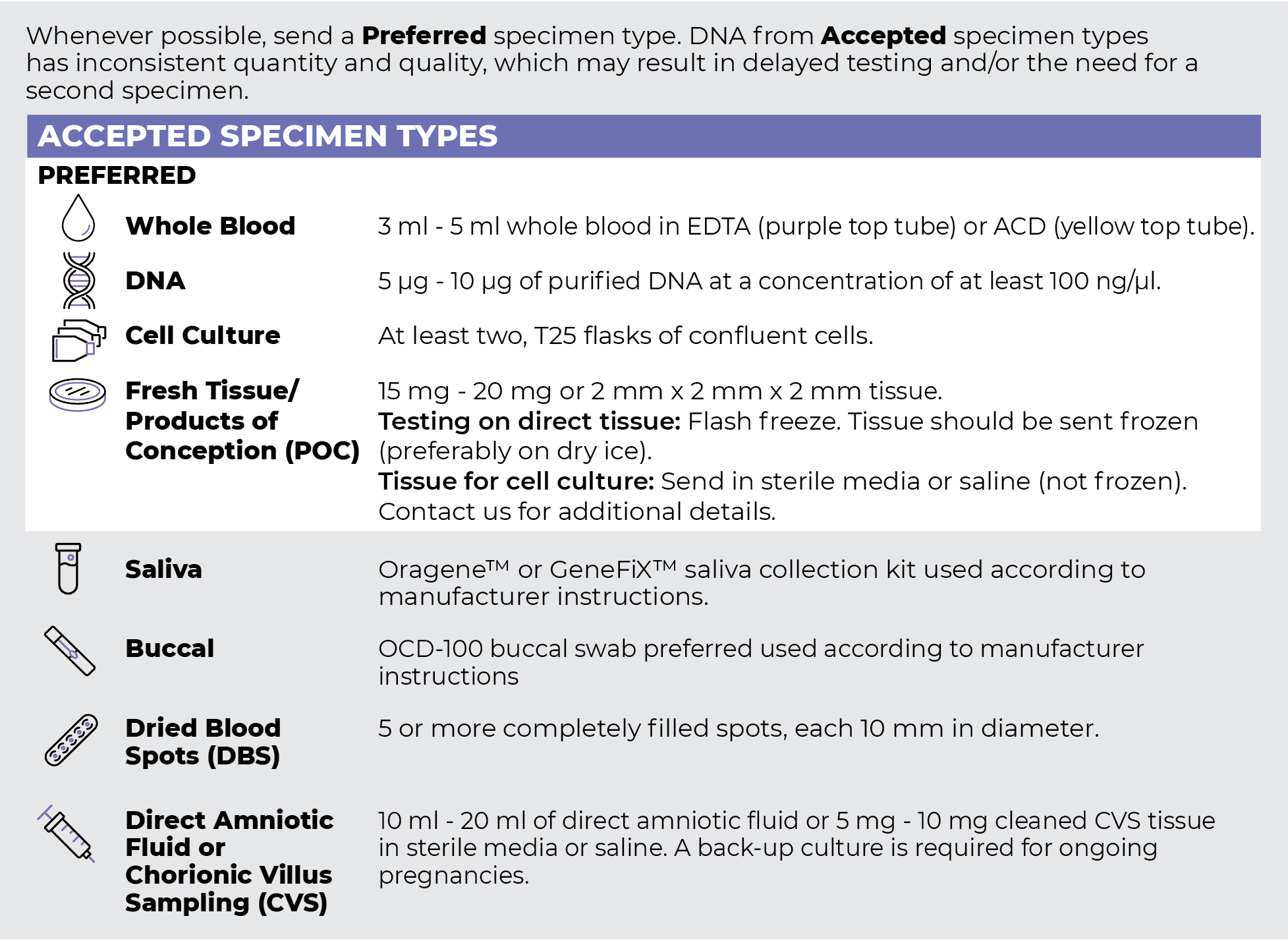
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.