



Cornelia de Lange Syndrome (CdLS) Panel
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Panel CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 10253 | Genes x (8) | 81479 | 81479(x16) | $990 | Order Options and Pricing |
Pricing Comments
We are happy to accommodate requests for testing single genes in this panel or a subset of these genes. The price will remain the list price. If desired, free reflex testing to remaining genes on panel is available. Alternatively, a single gene or subset of genes can also be ordered via our Custom Panel tool.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
Classic Cornelia de Lange syndrome (CdLS) is characterized by distinctive facial features, growth retardation, hirsutism, and upper limb reduction defects that range from subtle phalangeal abnormalities to oligodactyly. Craniofacial features include synophrys, arched eyebrows, long eyelashes, small upturned nose, small widely spaced teeth, and microcephaly. IQ ranges from below 30 to 102 (mean: 53). Many individuals demonstrate autistic and self-destructive tendencies. Frequent findings include cardiac septal defects, gastrointestinal dysfunction, hearing loss, myopia, and cryptorchidism or hypoplastic genitalia. Individuals with a milder phenotype have less severe growth, cognitive, and limb involvement, but often have facial features consistent with CdLS (Deardorff et al. 2012). CHOPS syndrome is characterized by cognitive impairment, coarse facies, heart defects, obesity, pulmonary involvement, short stature, and skeletal dysplasia (Izumi et al. 2015). CHOPS syndrome clinically overlaps with Cornelia de Lange syndrome. KBG syndrome is characterized by macrodontia of central upper incisors, mental retardation, dysmorphic facial features, short statue, skeletal anomalies and global developmental delay, seizures, and intellectual disability. KBG clinically overlaps with Cornelia de Lange syndrome (Ockeloen et al. 2015; Parenti et al. 2016).
Genetics
Cornelia de Lange syndrome is caused by pathogenic variants in 8 genes: NIPBL, SMC3, RAD21, SMC1A, HDAC8, KMT2A, AFF4, and ANKRD11. NIPBL, SMC3, RAD21, KMT2A, AFF4 and ANKRD11-related CdLS are all inherited in autosomal dominant manner, while SMC1A and HDAC8-related CdLS are inherited in an X-linked manner. Female carriers of SMC1A and HDAC8 pathogenic variants may show variable symptoms depending on random X-inactivation (Hoppman-Chaney et al. 2012; Mannini et al. 2013). The NIPBL, SMC3, RAD21, SMC1A, and HDAC8 genes encode regulatory or structural components of the cohesin complex, which play a key role involving sister chromatid cohesion, DNA repair, gene regulation, and genome stability (Barbero 2013). The KMT2A protein (Histone-lysine N-methyltransferase 2A) coded by exons 1 to 36 of the KMT2A gene on 11q23.3 is a member of H3K4-specific methyltransferases that plays an essential role in early development and hematopoiesis (Shen et al. 2014). The AFF4 protein coded by the AFF4 gene is a key component of the super elongation complex that regulates transcriptional processes during embryogenesis (Izumi et al. 2015).
Approximately 70% of all CdLS patients have germline pathogenic variants in one of five genes: NIPBL (60%), SMC1A (5%), HDAC8 (5%), and less than 1% in SMC3 and RAD21 (Mannini et al. 2013; Kaiser et al. 2014, Boyle et al. 2015). One study reported that NIPBL somatic mosaicism was found in 23% of buccal swab samples of clinically diagnosed CdLS patients, who did not have a detectable pathogenic variant in the five known CdLS genes in their blood sample (Huisman et al. 2013).
Approximately 360 unique NIPBL pathogenic variants have been reported. They are: missense (23%), nonsense (15%), splicing (17%), small del/ins (33%), gross del/ins (12%), and one complex arrangement. 99% of NIPBL pathogenic variants occur de novo. It has been suggested that milder forms of CdLS are more likely to be caused by missense mutation, and more severe forms by truncating mutations (Gillis et al. 2004; Boyle et al. 2015).
Approximately 30 unique HDAC8 pathogenic variants have been reported. They are: missense (67%), large del/ins (23%), nonsense (10%), splicing (6%), one small deletion (Human Gene Mutation Database; Kaiser et al. 2014).
Approximately 15 unique SMC3 pathogenic variants have been reported. They are: missense (60%), nonsense (1), in-frame small deletion (33%) and one nonsense. 9 out of those 15 pathogenic variants occur de novo, which explain 1%–2% of CdLS-like phenotypes (Gil-Rodríguez et al. 2015).
Approximately 50 unique SMC1A pathogenic variants have been reported. They are: missense (58%), small deletion (14%), large del/ins, and complex rearrangement (10%) and one splicing (Musio et al. 2006; Baquero-Montoya et al. 2014; Ansari et al. 2014). Less than 10 unique RAD21 pathogenic variants have been reported. They are: missense (2), splicing (1), small deletion (1), and large deletions (3) (Deardorff et al. 2012; Minor et al. 2014; Ansari et al. 2014).
KMT2A-related CdLS is inherited in autosomal dominant manner. Pathogenic KMT2A variants mainly cause Wiedemann-Steiner syndrome, which is characterized by hypertrichosis cubiti, short status, intellectual disability, and dysmorphic facial and skeletal features (Jones et al. 2012). To date, approximately 15 unique pathogenic variants in KMT2A have been documented; most pathogenic variants were found in patients with Wiedemann-Steiner syndrome and only one de novo truncating pathogenic variant was reported in one patient with CdLS. The pathogenic variants include missense (4), nonsense (4), small del/ins (5), splicing (1) and large deletion (1). De novo pathogenic variants were found in five of the six Wiedemann-Steiner syndrome cases (Jones et al. 2012; Human Gene Mutation Database).
AFF4-related CHOPS syndrome is inherited in an autosomal dominant manner. Only 3 unique de novo missense AFF4 variants were reported in 3 unrelated probands affected with CHOPS syndrome by exome sequencing (Izumi et al. 2015).
ANKRD11-related CdLS and KBG syndrome are inherited in an autosomal dominant manner. Almost 50 pathogenic variants were reported, mainly found in patients with KBG syndrome. Most of the variants are truncating (nonsense and small deletion/insertion). Missense and splice variants are rare. Large deletion/duplications account for ~30% of reported ANKRD11 pathogenic variants (HGMD). Only 4 unique, pathogenic variants were reported in patients affected with CdLS, they all de novo, truncating variants (nonsense and small deletion/duplication) (Ansari et al. 2014; Parenti I. et al. 2016).
Clinical Sensitivity - Sequencing with CNV PGxome
Over 70% of all CdLS patients harbor a pathogenic variant in NIPBL, SMC3, RAD21, SMC1A, or HDAC8 (Boyle et al. 2015). Only a few patients have been reported with pathogenic variants in AFF4, ANKRD11 and KMT2A.
Intragenic NIPBL deletions and a duplication were identified in 13 (2.5%) out of 510 CdLS cases (12 deletions and 1 duplication) (Cheng et al. 2014). Large deletions and duplications account for 11% of reported SMC1A pathogenic variants (Gilissen et al. 2014; Baquero-Montoya et al. 2014). There are no reported large deletions or duplications in SMC3 (Human Gene Mutation Database). Deletion and duplication testing is currently unavailable for the HDAC8 and RAD21 genes. Large deletion/duplication accounts for ~30% of reported pathogenic variants in ANKRD11 (Human Gene Mutation Database).
Testing Strategy
This test is performed using Next-Gen sequencing with additional Sanger sequencing as necessary.
This panel provides 100% coverage of all coding exons of the genes plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define coverage as ≥20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Candidates for this test are patients with clinical features consistent with Cornelia de Lange syndrome.
Candidates for this test are patients with clinical features consistent with Cornelia de Lange syndrome.
Genes
| Official Gene Symbol | OMIM ID |
|---|---|
| AFF4 | 604417 |
| ANKRD11 | 611192 |
| HDAC8 | 300269 |
| KMT2A | 159555 |
| NIPBL | 608667 |
| RAD21 | 606462 |
| SMC1A | 300040 |
| SMC3 | 606062 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
| Name | Inheritance | OMIM ID |
|---|---|---|
| CHOPS Syndrome | AD | 616368 |
| Cornelia de Lange syndrome 1 | AD | 122470 |
| Cornelia de Lange syndrome 2 | XL | 300590 |
| Cornelia de Lange syndrome 3 | AD | 610759 |
| Cornelia de Lange syndrome 4 | AD | 614701 |
| Cornelia de Lange syndrome 5 | XL | 300882 |
| KBG Syndrome | AD | 148050 |
| Wiedemann-Steiner Syndrome | AD | 605130 |
Related Test
| Name |
|---|
| PGxome® |
Citations
- Ansari M. et al. 2014. Journal of Medical Genetics. 51: 659-68. PubMed ID: 25125236
- Baquero-Montoya et al. 2014. PubMed ID: 23683030
- Barbero J.L. 2013. The Application of Clinical Genetics. 6: 15-23. PubMed ID: 23882154
- Boyle et al. 2015. PubMed ID: 25209348
- Cheng et al. 2014. PubMed ID: 24689074
- Deardorff M.A. et al. 2012. American Journal of Human Genetics. 90: 1014-27. PubMed ID: 22633399
- Gil-Rodríguez M.C. et al. 2015. Human Mutation. 36: 454-62. PubMed ID: 25655089
- Gilissen et al. 2014. PubMed ID: 24896178
- Gillis L.A. et al. 2004. American Journal of Human Genetics. 75: 610-23. PubMed ID: 15318302
- Hoppman-Chaney N. et al. 2012. American Journal of Medical Genetics. Part A. 158A: 193-8. PubMed ID: 22106055
- Huisman S.A. et al. 2013. Journal of Medical Genetics. 50: 339-44. PubMed ID: 23505322
- Human Gene Mutation Database (Bio-base).
- Izumi K. et al. 2015. Nature Genetics. 47: 338-44. PubMed ID: 25730767
- Jones et al. 2012. PubMed ID: 22795537
- Kaiser F.J. et al. 2014. Human Molecular Genetics. 23: 2888-900. PubMed ID: 24403048
- Mannini L. et al. 2013. Human Mutation. 34: 1589-96. PubMed ID: 24038889
- Minor A. et al. 2014. Gene. 537: 279-84. PubMed ID: 24378232
- Musio A. et al. 2006. Nature Genetics. 38: 528-30. PubMed ID: 16604071
- Ockeloen C.W. et al. 2015. European Journal of Human Genetics. 23: 1270. PubMed ID: 26269249
- Parenti I. et al. 2016. Clinical Genetics. 89: 74-81. PubMed ID: 25652421
- Shen E. et al. 2014. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences. 369: PubMed ID: 25135975
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
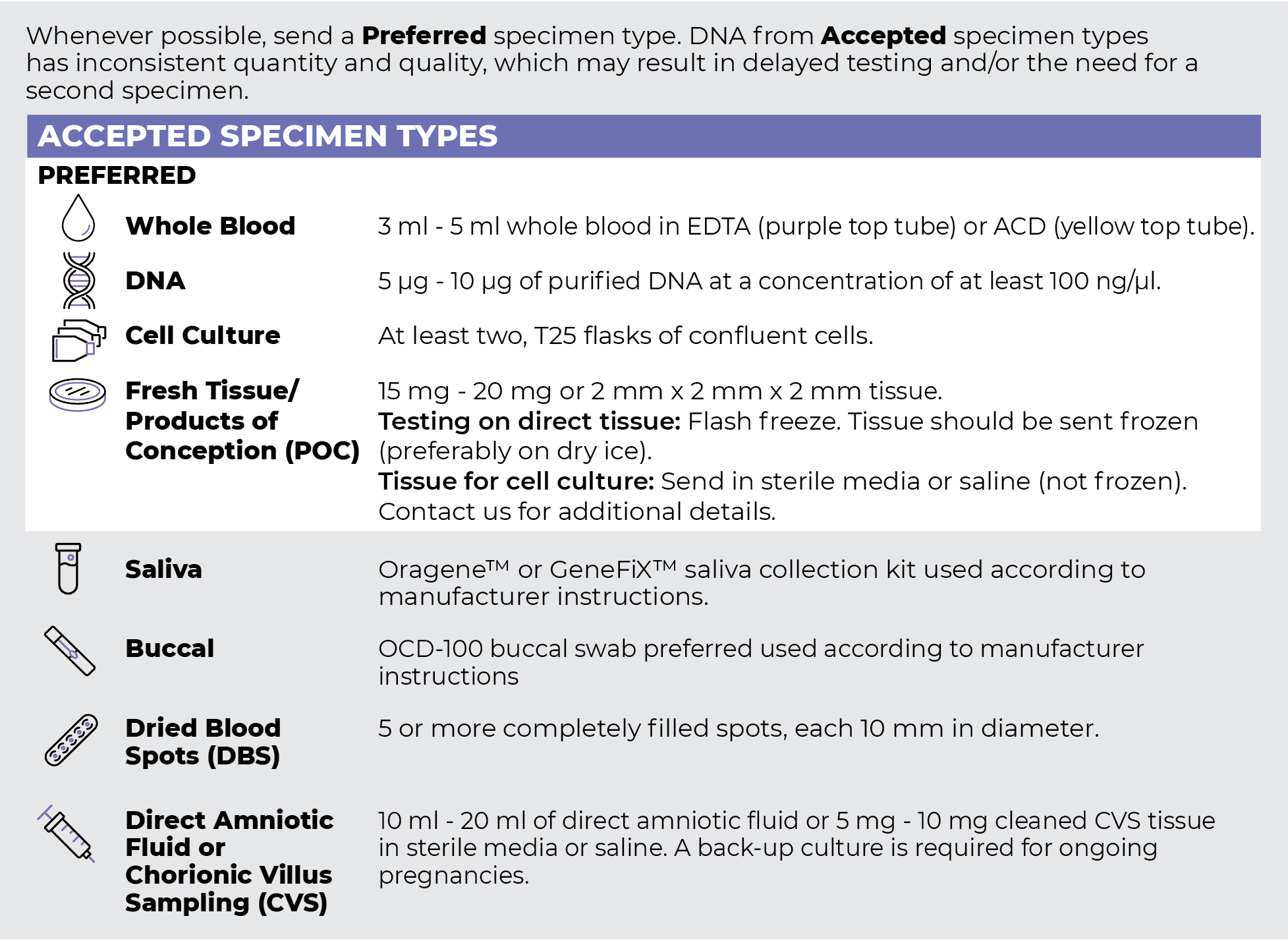
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.