



Hyperglycemia and Hypoglycemia via the GCK Gene
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Test CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 8427 | GCK | 81406 | 81406,81479 | $990 | Order Options and Pricing |
Pricing Comments
Our favored testing approach is exome based NextGen sequencing with CNV analysis. This will allow cost effective reflexing to PGxome or other exome based tests. However, if full gene Sanger sequencing is desired for STAT turnaround time, insurance, or other reasons, please see link below for Test Code, pricing, and turnaround time information. If the Sanger option is selected, CNV detection may be ordered through Test #600.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
The Sanger Sequencing method for this test is NY State approved.
For Sanger Sequencing click here.Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
Defects of glucokinase (hexokinase 4) can cause either hyperglycemia or hypoglycemia depending on underlying mutations of its encoding gene GCK (Osbak et al. Hum Mutat 30(11):1512-1526, 2009). Inactivating (loss-of-function) GCK mutations lead to two types of hyperglycemia: maturity-onset diabetes of the young (MODY) and permanent neonatal diabetes mellitus (PNDM). On the other hand, activating (gain-of-function) GCK mutations have been associated with hyperinsulinemic hypoglycemia, also known as congenital hyperinsulinism (CHI).
MODY is the most common type of monogenic diabetes, accounting for up to 5% of young adults diagnosed with diabetes (Owen. Clin Med 13(3):278-281, 2013; McDonald et al. Ann Clin Biochem 50(Pt 5):403-415, 2013). MODY typically presents in lean young adults before 25 years with an autosomal dominant family history. MODY patients have continuous production of endogenous insulin, absence of beta-cell autoimmunity and absence of signs of insulin resistance. As this non-insulin dependent type of diabetes is frequently misdiagnosed as Type 1 or Type 2 diabetes, a timely and accurate molecular diagnosis of MODY is essential to treatment decisions, prognosis, family screening and obstetric management of gestational diabetes (Pihoker et al. J Clin Endocrinol Metab 98(10):405540-62, 2013; Ellard et al. Diabetologia 51(4):546-553, 2008). Mutations in the GCK, HNF1A and HNF4A genes account for up to 80% of all MODY cases. GCK-MODY (MODY, type II or MODY2) represents a milder form of disease with a mild, asymptomatic and non-progressive fasting hyperglycaemia from birth without the requirement of pharmacological treatment. GCK-MODY is the most common form of MODY in children, accounting for about 30% of total MODY cases in the UK (Owen, 2013).
Neonatal diabetes mellitus, another type of monogenic diabetes, is defined as insulin-requiring hyperglycemia within the first 6 months (or even beyond 6 months) of life and is often associated with intrauterine growth retardation (Njølstad et al. N Engl J Med 344(21):1588-1592, 2001; Njølstad et al. Diabetes 52(11):2854-2860, 2003; Massa et al. Hum Mutat 25(1):22-27, 2005; Colombo et al. J Clin Invest. 118(6):2148-2156, 2008). About half of affected neonates have transient diabetes while the rest have permanent neonatal diabetes mellitus (PNDM; OMIM# 606176). Some severe PNDM patients may have neurological complications including developmental delay, epilepsy and neonatal diabetes (also termed DEND syndrome) (Molven et al. Expert Rev Mol Diagn 11(3):313-320, 2011). GCK-PNDM is caused by complete deficiency of glucokinase and thus represents the severe end of GCK- associated hyperglycemia.
Congenital hyperinsulinism (CHI) is a clinically and genetically heterogeneous condition characterized by hypoglycemia (Glaser et al. GeneReviews, 2003; Arnoux et al. Early Hum Dev 86(5):287-294, 2010). The age of disease onset ranges from the neonatal period with severe forms to infancy or childhood with milder forms. Severe patients typically have extremely low serum glucose while milder cases present with variable hypoglycemia. Affected newborns also develop nonspecific symptoms including seizures, apnea, hypotonia, and poor feeding. Severity of disease manifestations can vary within the same family.
Genetics
The GCK gene has 10 coding exons that encode glucokinase (hexokinase 4), which phosphorylates glucose to produce glucose-6-phosphate in glucose metabolism pathways (Osbak et al., 2009). Playing a crucial role in the regulation of insulin secretion, glucokinase has been termed the glucose sensor in pancreatic beta-cells. GCK defects throughout the whole gene include missense, nonsense, regulatory, splicing site mutations, small deletion/insertions while intragenic exon-level deletions/duplications are rare (Human Gene Mutation Database).
MODY is inherited in an autosomal dominant manner. So far, there are about 12 different genes associated with MODY-like phenotypes (Molven et al., 2011). Alongside HNF1A, HNF4A and HNF1B, GCK is among the most frequently involved genes in MODY (Owen, 2013). Approximately 85% of documented pathogenic GCK mutations are associated with MODY (Human Gene Mutation Database). Among these MODY-associated GCK mutations, about 60% are missense changes. Partial or whole gene deletions have been found to be rare in GCK-MODY (Ellard et al. Diabetologia 50(11):2313-2317, 2007; Garin et al. Clin Endocrinol 68(6):873-878, 2008). Phenotypes are notably similar for all MODY-associated GCK mutations (Ellard et al., 2008).
PNDM is commonly caused by mutations in the genes ABCC8 and KCNJ11, which encode the subunits of the beta-cell ATP-sensitive potassium (KATP) channel (Molven et al., 2011). Another relatively frequent cause of PNDM are mutations in the insulin gene (INS). Defects of GCK represent a rare cause of PNDM. GCK-PNDM is inherited recessively and only limited cases have been reported. PNDM-associated GCK defects include missense, nonsense and frameshift mutations, either homozygous or compound heterozygous which result in complete deficiency of glucokinase activity (Osbak et al., 2009).
CHI is genetically caused by defects in genes involved in regulation of insulin secretion from pancreatic beta-cells (Kapoor et al. Eur J Endocrinol 168(4):557-564, 2013; Snider et al. J Clin Endocrinol Metab 98(2):E355-363, 2013). GCK-related congenital hyperinsulinism (OMIM# 602485) is inherited in an autosomal dominant manner. Dominant activating GCK mutations are the third most common genetic cause of CHI (Snider et al., 2013; Glaser et al. N Engl J Med 338(4):226-230, 1998). GCK mutations have been found in both diazoxide-responsive (Christesen et al. Eur J Endocrinol 159(1):27-34, 2008) and diazoxide-unresponsive CHI patients (Snider et al., 2013). De novo GCK mutations in CHI are very common. All germline GCK mutations found in CHI patients are missense substitutions with a clustering at a confined region termed the allosteric activator site, which is remote to the substrate-binding site. Notably, a postzygotic mosaic GCK mutation (c.1361_1363dupCGG/p.Ala454dup) has been found in a patient’s pancreatic, but not peripheral blood, DNA (Snider et al., 2013). Eight other genes have also been associated with CHI including two KATP genes ABCC8 and KCNJ11, GLUD1, HADH, SLC16A1, HNF4A, HNF1A and UCP2 (Glaser et al., 2003; Kapoor et al.; Snider et al., 2013).
Clinical Sensitivity - Sequencing with CNV PGxome
The GCK mutation detection rate is widely variable depending on the age and clinical selection criteria of the target population (Osbak et al. Hum Mutat 30(11):1512-1526, 2009; Pihoker et al. J Clin Endocrinol Metab 98(10):405540-62, 2013). In a study of 82 children with incidental hyperglycaemia, 43% were found to carry GCK mutations (Feigerlová et al. Eur J Pediatr 165(7):446-452, 2006). The GCK mutation detection rate in a large cohort of patients with PNDM is unavailable because mutations have been only reported in limited individual cases. Defects of GCK represent a rare cause of PNDM. In a cohort of 298 diazoxide-nonresponsive CHI patients studied at the Hyperinsulinism Center in The Children’s Hospital of Philadelphia (CHOP), approximately 2% (7 of 298) were identified to have GCK mutations via DNA sequencing (Snider et al. J Clin Endocrinol Metab. 98(2):E355-63, 2013). In another cohort of 300 CHI patients studied in the United Kingdom, no GCK mutations were found in 105 diazoxide-nonresponsive or 183 diazoxide-responsive patients (Kapoor et al. Eur J Endocrinol 168(4):557-564, 2013). In a study of CHI patients enrolled from three European referral centers, the overall prevalence of GCK-related non-syndromic CHI was approximately 1%. In the medically responsive group, the prevalence of GCK-caused CHI was approximately 7% (Christesen et al. Eur J Endocrinol 159(1):27-34, 2008).
Testing Strategy
This test provides full coverage of all coding exons of the GCK gene plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define full coverage as >20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
It also includes targeted testing of a promoter regulatory mutation c.-557G>C (Gasperíková et al. Diabetes 58(8):1929-1935, 2009).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Candidates for this test include:
1) Patients with mild fasting hypoglycemia and pregnant women with gestational diabetes (Ellard et al., 2008).
2) Infants with neonatal diabetes, especially if some first degree relatives have a mild form of diabetes or glucose intolerance (Molven et al., 2011; Njølstad et al., 2001).
3) Patients with CHI, especially when ABCC8 and KCNJ11 are normal (Snider et al., 2013; Christesen et al., 2008).
Additionally, testing is indicated for family members of patients who have known GCK mutations. This test may also be considered for the reproductive partners of individuals who carry pathogenic variants in GCK.
Candidates for this test include:
1) Patients with mild fasting hypoglycemia and pregnant women with gestational diabetes (Ellard et al., 2008).
2) Infants with neonatal diabetes, especially if some first degree relatives have a mild form of diabetes or glucose intolerance (Molven et al., 2011; Njølstad et al., 2001).
3) Patients with CHI, especially when ABCC8 and KCNJ11 are normal (Snider et al., 2013; Christesen et al., 2008).
Additionally, testing is indicated for family members of patients who have known GCK mutations. This test may also be considered for the reproductive partners of individuals who carry pathogenic variants in GCK.
Gene
| Official Gene Symbol | OMIM ID |
|---|---|
| GCK | 138079 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
| Name | Inheritance | OMIM ID |
|---|---|---|
| Hyperinsulinemic Hypoglycemia, Familial 3 | AD | 602485 |
| Maturity-Onset Diabetes Of The Young, Type 2 | AD | 125851 |
| Permanent Neonatal Diabetes Mellitus | AR, AD | 606176 |
Citations
- Arnoux J.B. et al. 2010. Early Human Development. 86: 287-94. PubMed ID: 20550977
- Christesen, H. et al. (2008). “Activating glucokinase (GCK) mutations as a cause of medically responsive congenital hyperinsulinism: prevalence in children and characterisation of a novel GCK mutation.” Eur J Endocrinol 159(1):27-34. PubMed ID: 18450771
- Colombo, C. et al. (2008). “Seven mutations in the human insulin gene linked to permanent neonatal/infancy-onset diabetes mellitus.” J Clin Invest 118(6):2148-2156. PubMed ID: 18451997
- Ellard, S. et al. (2007). "Partial and whole gene deletion mutations of the GCK and HNF1A genes in maturity-onset diabetes of the young." Diabetologia 50(11):2313-2317. PubMed ID: 17828387
- Ellard, S. et al. (2008). "Best practice guidelines for the molecular genetic diagnosis of maturity-onset diabetes of the young." Diabetologia 51(4):546-553. PubMed ID: 18297260
- Feigerlová, E. et al. (2006). “Aetiological heterogeneity of asymptomatic hyperglycaemia in children and adolescents.” Eur J Pediatr 165(7):446-452. PubMed ID: 16602010
- Garin, I. et al. (2008). "Haploinsufficiency at GCK gene is not a frequent event in MODY2 patients." Clin Endocrinol 68(6):873-878. PubMed ID: 18248649
- Gasperíková, D. et al. (2009). "Identification of a novel beta-cell glucokinase (GCK) promoter mutation (-71G>C) that modulates GCK gene expression through loss of allele-specific Sp1 binding causing mild fasting hyperglycemia in humans." Diabetes 58(8):1929-1935. PubMed ID: 19411616
- Glaser B. 2003. Familial Hyperinsulinism. In: Pagon RA, Adam MP, Ardinger HH, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301549
- Glaser, B. et al. (1998). "Familial hyperinsulinism caused by an activating glucokinase mutation." N Engl J Med 338(4):226-230. PubMed ID: 9435328
- Human Gene Mutation Database (Bio-base).
- Kapoor R.R. et al. 2013. European Journal of Endocrinology / European Federation of Endocrine Societies. 168: 557-64. PubMed ID: 23345197
- Massa, O. et al. (2005). “KCNJ11 activating mutations in Italian patients with permanent neonatal diabetes.” Hum Mutat 25(1):22-27. PubMed ID: 15580558
- McDonald, T. et al. (2013). “Maturity onset diabetes of the young: identification and diagnosis.” Ann Clin Biochem 50(Pt 5):403-415. PubMed ID: 23878349
- Molven, A. et al. (2011). "Role of molecular genetics in transforming diagnosis of diabetes mellitus." Expert Rev Mol Diagn 11(3):313-320. PubMed ID: 21463240
- Njølstad, P. et al. (2001). "Neonatal diabetes mellitus due to complete glucokinase deficiency." N Engl J Med 344(21):1588-1592. PubMed ID: 11372010
- Njølstad, P. et al. (2003). “Permanent neonatal diabetes caused by glucokinase deficiency: inborn error of the glucose-insulin signaling pathway.” Diabetes 52(11):2854-2860. PubMed ID: 14578306
- Osbak, K. et al. (2009). "Update on mutations in glucokinase (GCK), which cause maturity-onset diabetes of the young, permanent neonatal diabetes, and hyperinsulinemic hypoglycemia." Hum Mutat 30(11):1512-1526. PubMed ID: 19790256
- Owen, K. (2013). "Monogenic diabetes: old and new approaches to diagnosis." Clin Med 13(3):278-281. PubMed ID: 23760703
- Pihoker C, Gilliam LK, Ellard S, Dabelea D, Davis C, Dolan LM, Greenbaum CJ, Imperatore G, Lawrence JM, Marcovina SM, Mayer-Davis E, Rodriguez BL, et al. 2013. Prevalence, characteristics and clinical diagnosis of maturity onset diabetes of the young due to mutations in HNF1A, HNF4A, and glucokinase: results from the SEARCH for Diabetes in Youth. J. Clin. Endocrinol. Metab. 98: 4055–4062. PubMed ID: 23771925
- Snider K.E. et al. 2013. The Journal of Clinical Endocrinology and Metabolism. 98: E355-63. PubMed ID: 23275527
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
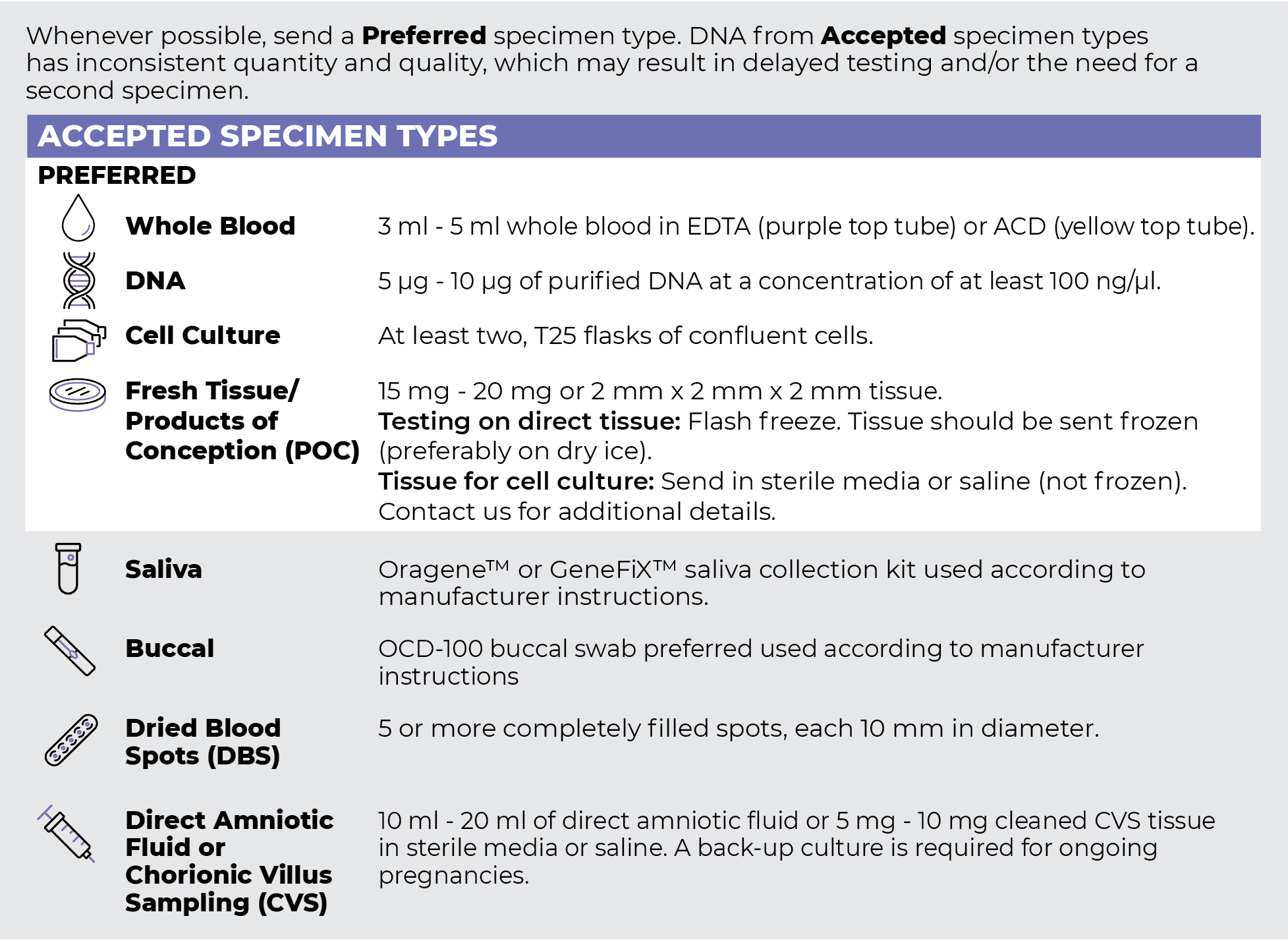
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.