



Charcot-Marie-Tooth disease, axonal, type 20, Spinal muscular atrophy with lower extremity predominance and Mental retardation, autosomal dominant type 13 via the DYNC1H1 Gene
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Test CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 11273 | DYNC1H1 | 81479 | 81479,81479 | $990 | Order Options and Pricing |
Pricing Comments
Our favored testing approach is exome based NextGen sequencing with CNV analysis. This will allow cost effective reflexing to PGxome or other exome based tests. However, if full gene Sanger sequencing is desired for STAT turnaround time, insurance, or other reasons, please see link below for Test Code, pricing, and turnaround time information.
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
The Sanger Sequencing method for this test is NY State approved.
For Sanger Sequencing click here.Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
In a large four generation pedigree affected with type 2 Charcot-Marie-Tooth disease (OMIM 614228) a heterozygous DYNC1H1 missense mutation was found to cosegregate with disease (Weedon et al. Am J Hum Genet 89:308-312, 2011). Some patients presented at birth with club foot or pes cavus and others were reported to require multiple surgeries to correct foot and ankle problems. Other individuals presented early in the second decade of life with abnormal gait, difficulty running, and frequent falls. Still others presented with delayed motor milestones, developmental delays, or speech delays. Common clinical features include proximal and distal lower limb weakness and wasting. Reflexes were reduced in some, but not all affected individuals.
Two cases of autosomal dominant mental retardation have been attributed to de novo mutations in the DYNC1H1 gene (OMIM 614563). Both cases were discovered by family-based exome sequencing. The first affected individual had, in addition to severe intellectual disability, hypotonia, hyporeflexia, mild dysmorphic facial features, and broad-based waddling gait with toe-walking (Vissers et al. Nat Genet 42:1109-1112, 2010). The patient was also found to have deficient gyration of the frontal lobes by brain MRI studies. The second case was ascertained at the age of 51years (Willemsen et al. J Med Genet 49:179-183, 2012). This patient also had severe intellectual impairment since infancy and was unable to walk or speak. Additionally, she was mildly dysmorphic and had abnormal cortex structure with deficient gyration.
In a large six generation pedigree affected with spinal muscular atrophy with lower extremity predominance (SMALED, OMIM 158600) a heterozygous DYNC1H1 missense mutation was found to cosegregate with disease (Harms et al. Neurology 78:1714-1720, 2012). In affected individuals from this pedigree muscle atrophy and weakness was confined to the lower limbs and there was little progression. Early clinical signs included a waddling gait and awkward running. Deep tendon reflexes were reduced at the knees and nerve conduction studies were consistent with motor neuron disease without sensory involvement. Thirty-two other patients with SMALED were tested by DYNC1H1 sequencing and two were found to have missense mutations. Notably, all three SMALED-causing mutations are located in the tail domain of the DYNC1H1 protein.
Genetics
DYNC1H1-related disorders are inherited in an autosomal dominant mode. Individuals with Charcot-Marie-Tooth disease type 20 spinal muscular atrophy with lower extremity predominance inherit a DYNC1H1 mutation from an affected parent. Mutations resulting in autosomal dominant mental retardation type 13 have been shown to be de novo (Vissers et al. Nature Genet 42: 1109-1112, 2010; Willemsen et al. J Med Genet 49:179-183, 2012). Missense mutations are the only form of pathogenic gene variant thus far reported for the DYNC1H1 gene.
Clinical Sensitivity - Sequencing with CNV PGxome
Analytical sensitivity may be high because all DYNC1H1 mutations reported to date are expected to be detectable by direct sequencing of genomic DNA. Clinical sensitivity is problematic to predict due to genetic heterogeneity of these disorders and the death of documented cases. Vissers et al. (Nat Genet 42: 1109-1112, 2010) tested exomes of ten patients with unexplained severe intellectual impairment and found one to have a de novo DYNC1H1 mutation. Harms et al. (Neurology 78: 1714-1720, 2012) found two patients with DYNC1H1 mutations among 32 probands with spinal muscular atrophy with lower extremity predominance.
Testing Strategy
This test provides full coverage of all coding exons of the DYNC1H1 gene plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define full coverage as >20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Patients with clinical features of one of the DYNC1H1-related disorders.
Patients with clinical features of one of the DYNC1H1-related disorders.
Gene
| Official Gene Symbol | OMIM ID |
|---|---|
| DYNC1H1 | 600112 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Diseases
Citations
- Harms et al. Mutations in the tail domain of DYNC1H1 cause dominant spinal muscular atrophy.Neurology 78: 1714-1720, 2012. PubMed ID: 22459677
- Vissers et al. A de novo paradigm for mental retardation. Nature Genet. 42: 1109-1112, 2010. PubMed ID: 21076407
- Weedon et al. Exome sequencing identifies a DYNC1H1 mutation in a large pedigree with dominant axonal Charcot-Marie-Tooth disease. Am J Hum Genet 89: 308-312, 2011. PubMed ID: 21820100
- Willemsen et al. Mutations in DYNC1H1 cause severe intellectual disability with neuronal migration defects. J Med Genet 49: 179-183, 2012. PubMed ID: 22368300
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
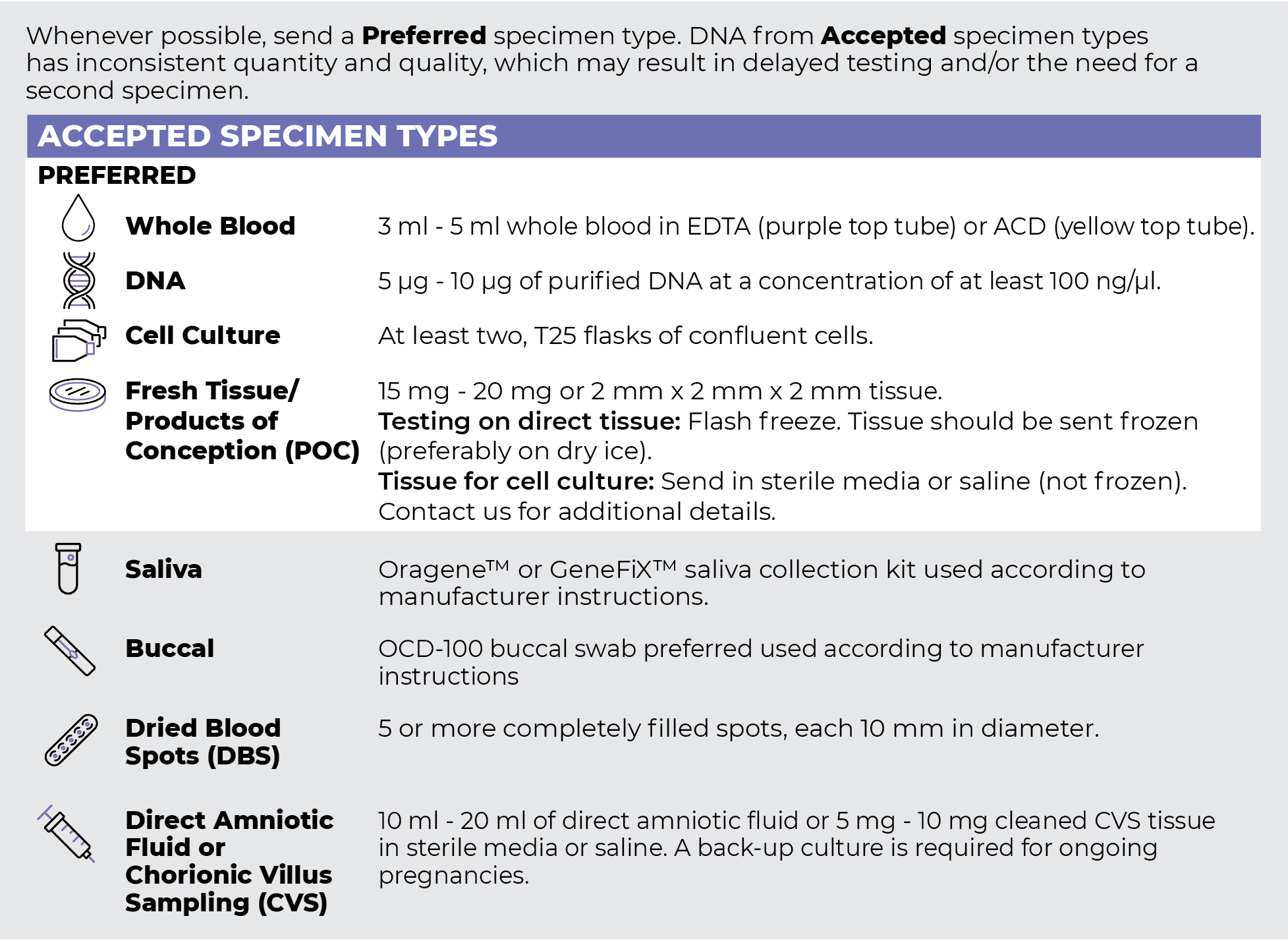
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.