



CHARGE and Kallmann Syndromes via the CHD7 Gene
Summary and Pricing 
Test Method
Exome Sequencing with CNV Detection| Test Code | Test Copy Genes | Test CPT Code | Gene CPT Codes Copy CPT Code | Base Price | |
|---|---|---|---|---|---|
| 4633 | CHD7 | 81407 | 81407,81479 | $990 | Order Options and Pricing |
An additional 25% charge will be applied to STAT orders. STAT orders are prioritized throughout the testing process.
Click here for costs to reflex to whole PGxome (if original test is on PGxome Sequencing platform).
Click here for costs to reflex to whole PGnome (if original test is on PGnome Sequencing platform).
Turnaround Time
3 weeks on average for standard orders or 2 weeks on average for STAT orders.
Please note: Once the testing process begins, an Estimated Report Date (ERD) range will be displayed in the portal. This is the most accurate prediction of when your report will be complete and may differ from the average TAT published on our website. About 85% of our tests will be reported within or before the ERD range. We will notify you of significant delays or holds which will impact the ERD. Learn more about turnaround times here.
Targeted Testing
For ordering sequencing of targeted known variants, go to our Targeted Variants page.
Clinical Features and Genetics
Clinical Features
CHARGE syndrome is a severe developmental disorder characterized by multiple congenital defects involving sensory and mediastinal organs. It is a clinically heterogeneous disorder in regards to symptoms and severity. Hallmark features include ocular coloboma; choanal atresia; cranial nerve abnormalities leading to facial palsy, loss of sense of smell, feeding, swallowing and breathing difficulties; and external and inner ear malformations resulting in hearing loss and reduced sense of balance. Additional features include hypogonadotropic hypogonadism, which manifests as incomplete or absent puberty and infertility; genital hypoplasia; growth and developmental delay; a wide variety of heart defects; cleft lip or palate; and distinctive facial features. CHARGE syndrome is usually diagnosed during childhood. Diagnosis is made based on the presence of a combination of major and minor clinical features (Blake et al. 1998; Verloes et al. 2005). Magnetic resonance imaging (MRI) of the temporal bones reveals abnormalities in the semicircular canal (Amiel et al 2001). In rare cases, CHARGE syndrome has been detected in adult individuals only after the birth of a child with the major characteristic features of the disease (Hughes et al. 2014). It has also been diagnosed antenatally (Legendre et al. 2012). CHARGE syndrome affects individuals worldwide with an incidence of approximately 1 case in 12,500 live births (Källén et al. 1999). Higher incidences have been reported in the Atlantic provinces of Newfoundland and Labrador and the Maritime Provinces (Issekutz et al. 2005). See also (Lalani et al. 2012) and the CHARGE Syndrome Foundation (http://www.chargesyndrome.org/foundation.asp).
Kallmann syndrome has clinical overlap with CHARGE syndrome. It is characterized by hypogonadotropic hypogonadism and impaired sense of smell as the result of deficient hypothalamic gonadotropin-releasing hormone and agenesis of the olfactory lobes, respectively. Additional features include unilateral failure of kidney development; abnormalities in tooth development; cleft lip and/or palate; and bimanual synkinesis, which is manifested by involuntary movements of one hand that mimic the other hand (Dode et al. 2003; Kaplan et al. 2010).
Genetics
CHARGE and Kallmann syndromes are autosomal dominant conditions. More than 95% of patients with a clinical diagnosis of CHARGE syndrome based on the Blake or Verloes criteria have heterozygous mutations in the CHD7 gene (Blake 1998; Verloes, 2005; Blake 2011). Over 680 different causative mutations, located throughout the length of the gene, are listed in public databases (Human Gene Mutation Database; CHD7 Mutation Database). The great majority result in truncated proteins, and include nonsense, splicing, small deletions and insertions. About 2% of all pathogenic variants are large deletions, some of which include the entire CHD7 gene (Wincent et al. 2008). Chromosomal abnormalities as the result of balanced translocations, rearrangements, or interstitial deletions have been reported (Hurst et al. 1991; Johnson 2005; Arrington et al. 2005). Although most disease-causing variants are de novo, familial cases have been reported (Jongmans et al. 2008; Hughes et al. 2014). In these families, clinical features are usually variable among affected individuals and may be very mild. Parental mosaicism, both somatic and germline, have been detected (Jongmans et al 2006; Pauli et al. 2009).
About 30 CHD7 pathogenic variants are reported in patients with KS; they account for ~ 11% of patients with a clinical diagnosis (Marcos et al. 2014). Unlike CHARGE-causative variants, the majority of KS-causative variants are missense. To date, no large deletions, duplications, or complex rearrangements were reported. Most cases are sporadic.
The CHD7 gene encodes the chromodomain helicase DNA-binding protein 7 that is required for normal mammalian development.
Clinical Sensitivity - Sequencing with CNV PGxome
The sensitivity of this test varies based on the criteria used for diagnosis. Mutations in CHD7 are detected in over 95% of patients with a clinical diagnosis based on Blake or Verloes criteria (Blake et al. 1998; Verloes et al. 2005). CHD7 mutations are found in 60-70% of patients who are suspected to have CHARGE syndrome (Blake et al. 2011). About 11% of patients with a clinical diagnosis of Kallmann syndrome have pathogenic variants in CHD7 (Marcos et al. 2014).
Large pathogenic deletions have been reported in less than 5% of patients with a clinical diagnosis of CHARGE syndrome (Bergman et al. 2008; Wincent et al. 2009; Blake et al. 2011).
Testing Strategy
This test provides full coverage of all coding exons of the CHD7 gene plus 10 bases of flanking noncoding DNA in all available transcripts along with other non-coding regions in which pathogenic variants have been identified at PreventionGenetics or reported elsewhere. We define full coverage as >20X NGS reads or Sanger sequencing. PGnome panels typically provide slightly increased coverage over the PGxome equivalent. PGnome sequencing panels have the added benefit of additional analysis and reporting of deep intronic regions (where applicable).
Dependent on the sequencing backbone selected for this testing, discounted reflex testing to any other similar backbone-based test is available (i.e., PGxome panel to whole PGxome; PGnome panel to whole PGnome).
Indications for Test
Patients presenting with the major clinical criteria or a combination of minor and major criteria for CHARGE syndrome as described (Blake et al. 1998; Verloes et al. 2005). For Kallmann syndrome, hypogonadotropic hypogonadism and impaired sense of smell.
Patients presenting with the major clinical criteria or a combination of minor and major criteria for CHARGE syndrome as described (Blake et al. 1998; Verloes et al. 2005). For Kallmann syndrome, hypogonadotropic hypogonadism and impaired sense of smell.
Gene
| Official Gene Symbol | OMIM ID |
|---|---|
| CHD7 | 608892 |
| Inheritance | Abbreviation |
|---|---|
| Autosomal Dominant | AD |
| Autosomal Recessive | AR |
| X-Linked | XL |
| Mitochondrial | MT |
Disease
| Name | Inheritance | OMIM ID |
|---|---|---|
| CHARGE Association | AD | 214800 |
Related Tests
| Name |
|---|
| CHARGE and Kallmann Syndromes Panel |
| Hypogonadotropic Hypogonadism/Kallmann Syndrome Panel |
| Kallmann Syndrome (KS) Panel |
Citations
- Amiel J, Attieé-Bitach T, Marianowski R, Cormier-Daire V, Abadie V, Bonnet D, Gonzales M, Chemouny S, Brunelle F, Munnich A, Manach Y, Lyonnet S. 2001. Temporal bone anomaly proposed as a major criteria for diagnosis of CHARGE syndrome. Am. J. Med. Genet. 99: 124–127. PubMed ID: 11241470
- Arrington CB, Cowley BC, Nightingale DR, Zhou H, Brothman AR, Viskochil DH. 2005. Interstitial deletion 8q11.2-q13 with congenital anomalies of CHARGE association. American Journal of Medical Genetics Part A 133A: 326–330. PubMed ID: 15672384
- Bergman JEH, Wijs I de, Jongmans MCJ, Admiraal RJ, Hoefsloot LH, Ravenswaaij-Arts CMA van. 2008. Exon copy number alterations of the CHD7 gene are not a major cause of CHARGE and CHARGE-like syndrome. Eur J Med Genet 51: 417–425. PubMed ID: 18472328
- Blake K, Ravenswaaij-Arts CM van, Hoefsloot L, Verloes A. 2011. Clinical utility gene card for: CHARGE syndrome. European Journal of Human Genetics 19: PubMed ID: 21407266
- Blake KD, Davenport SL, Hall BD, Hefner MA, Pagon RA, Williams MS, Lin AE, Graham JM Jr. 1998. CHARGE association: an update and review for the primary pediatrician. Clin Pediatr (Phila) 37: 159–173. PubMed ID: 9545604
- CHARGE Syndrome Foundation
- CHD7 Mutation Database
- Dodé C, Levilliers J, Dupont J-M, Paepe AD, Dû NL, Soussi-Yanicostas N, Coimbra RS, Delmaghani S, Compain-Nouaille S, Baverel F, Pêcheux C, Tessier DL, et al. 2003. Loss-of-function mutations in FGFR1 cause autosomal dominant Kallmann syndrome. Nature Genetics 33: 463–465. PubMed ID: 12627230
- Hughes SS, Welsh HI, Safina NP, Bejaoui K, Ardinger HH. 2014. Family history and clefting as major criteria for CHARGE syndrome. American Journal of Medical Genetics Part A 164: 48–53. PubMed ID: 24214489
- Human Gene Mutation Database (Bio-base).
- Hurst JA, Meinecke P, Baraitser M. 1991. Balanced t(6;8)(6p8p;6q8q) and the CHARGE association. J. Med. Genet. 28: 54–55. PubMed ID: 1999835
- Issekutz KA, Graham JM, Prasad C, Smith IM, Blake KD. 2005. An epidemiological analysis of CHARGE syndrome: Preliminary results from a Canadian study. American Journal of Medical Genetics Part A 133A: 309–317. PubMed ID: 15637722
- Johnson D. 2005. Confirmation of CHD7 as a cause of CHARGE association identified by mapping a balanced chromosome translocation in affected monozygotic twins. Journal of Medical Genetics 43: 280–284. PubMed ID: 16118347
- Jongmans MCJ, Hoefsloot LH, Donk KP van der, Admiraal RJ, Magee A, Laar I van de, Hendriks Y, Verheij JBGM, Walpole I, Brunner HG, Ravenswaaij CMA van. 2008. Familial CHARGE syndrome and theCHD7 gene: A recurrent missense mutation, intrafamilial recurrence and variability. American Journal of Medical Genetics Part A 146A: 43–50. PubMed ID: 18074359
- Jongmans MCJ. 2005. CHARGE syndrome: the phenotypic spectrum of mutations in the CHD7 gene. Journal of Medical Genetics 43: 306–314. PubMed ID: 16155193
- Källén K, Robert E, Mastroiacovo P, Castilla EE, Källén B. 1999. CHARGE Association in newborns: a registry-based study. Teratology 60: 334–343. PubMed ID: 10590394
- Kaplan JD, Bernstein JA, Kwan A, Hudgins L. 2010. Clues to an early diagnosis of Kallmann syndrome. Am. J. Med. Genet. A 152A: 2796–2801. PubMed ID: 20949504
- Lalani SR, Hefner MA, Belmont JW, Davenport SL. 2012. CHARGE Syndrome. In: Pagon RA, Adam MP, Ardinger HH, Bird TD, Dolan CR, Fong C-T, Smith RJ, and Stephens K, editors. GeneReviews(®), Seattle (WA): University of Washington, Seattle. PubMed ID: 20301296
- Legendre M, Gonzales M, Goudefroye G, Bilan F, Parisot P, Perez M-J, Bonnière M, Bessières B, Martinovic J, Delezoide A-L, Jossic F, Fallet-Bianco C, Bucourt M, Tantau J, Loget P, Loeuillet L, Laurent N, Leroy B, Salhi H, Bigi N, Rouleau C, Guimiot F, Quélin C, Bazin A, Alby C, Ichkou A, Gesny R, Kitzis A, Ville Y, Lyonnet S, Razavi F, Gilbert-Dussardier B, Vekemans M, Attié-Bitach T. 2012. Antenatal spectrum of CHARGE syndrome in 40 fetuses with CHD7 mutations. J. Med. Genet. 49: 698–707. PubMed ID: 23024289
- Marcos S, Sarfati J, Leroy C, Fouveaut C, Parent P, Metz C, Wolczynski S, Gérard M, Bieth E, Kurtz F, Verier-Mine O, Perrin L, Archambeaud F, Cabrol S, Rodien P, Hove H, Prescott T, Lacombe D, Christin-Maitre S, Touraine P, Hieronimus S, Dewailly D, Young J, Pugeat M, Hardelin JP, Dodé C. 2014. The Prevalence of CHD7 Missense Versus Truncating Mutations Is Higher in Patients With Kallmann Syndrome Than in Typical CHARGE Patients. The Journal of Clinical Endocrinology & Metabolism 99: E2138–E2143. PubMed ID: 25077900
- Pauli S, Pieper L, Häberle J, Grzmil P, Burfeind P, Steckel M, Lenz U, Michelmann H. 2009. Proven germline mosaicism in a father of two children with CHARGE syndrome. Clinical Genetics 75: 473–479. PubMed ID: 19475719
- Verloes A. 2005. Updated diagnostic criteria for CHARGE syndrome: A proposal. American Journal of Medical Genetics Part A 133A: 306–308. PubMed ID: 15666308
- Vissers LELM, Ravenswaaij CMA van, Admiraal R, Hurst JA, Vries BBA de, Janssen IM, Vliet WA van der, Huys EHLPG, Jong PJ de, Hamel BCJ, Schoenmakers EFPM, Brunner HG, Veltman JA, van Kessel AG. 2004. Mutations in a new member of the chromodomain gene family cause CHARGE syndrome. Nature Genetics 36: 955–957. PubMed ID: 15300250
- Wincent J, Holmberg E, Strömland K, Soller M, Mirzaei L, Djureinovic T, Robinson K, Anderlid B, Schoumans J. 2008. CHD7 mutation spectrum in 28 Swedish patients diagnosed with CHARGE syndrome. Clinical Genetics 74: 31–38. PubMed ID: 18445044
- Wincent J, Schulze A, Schoumans J. 2009. Detection of CHD7 deletions by MLPA in CHARGE syndrome patients with a less typical phenotype. Eur J Med Genet 52: 271–272. PubMed ID: 19248844
Ordering/Specimens
Ordering Options
We offer several options when ordering sequencing tests. For more information on these options, see our Ordering Instructions page. To view available options, click on the Order Options button within the test description.
myPrevent - Online Ordering
- The test can be added to your online orders in the Summary and Pricing section.
- Once the test has been added log in to myPrevent to fill out an online requisition form.
- PGnome sequencing panels can be ordered via the myPrevent portal only at this time.
Requisition Form
- A completed requisition form must accompany all specimens.
- Billing information along with specimen and shipping instructions are within the requisition form.
- All testing must be ordered by a qualified healthcare provider.
For Requisition Forms, visit our Forms page
If ordering a Duo or Trio test, the proband and all comparator samples are required to initiate testing. If we do not receive all required samples for the test ordered within 21 days, we will convert the order to the most effective testing strategy with the samples available. Prior authorization and/or billing in place may be impacted by a change in test code.
Specimen Types
Specimen Requirements and Shipping Details
PGxome (Exome) Sequencing Panel
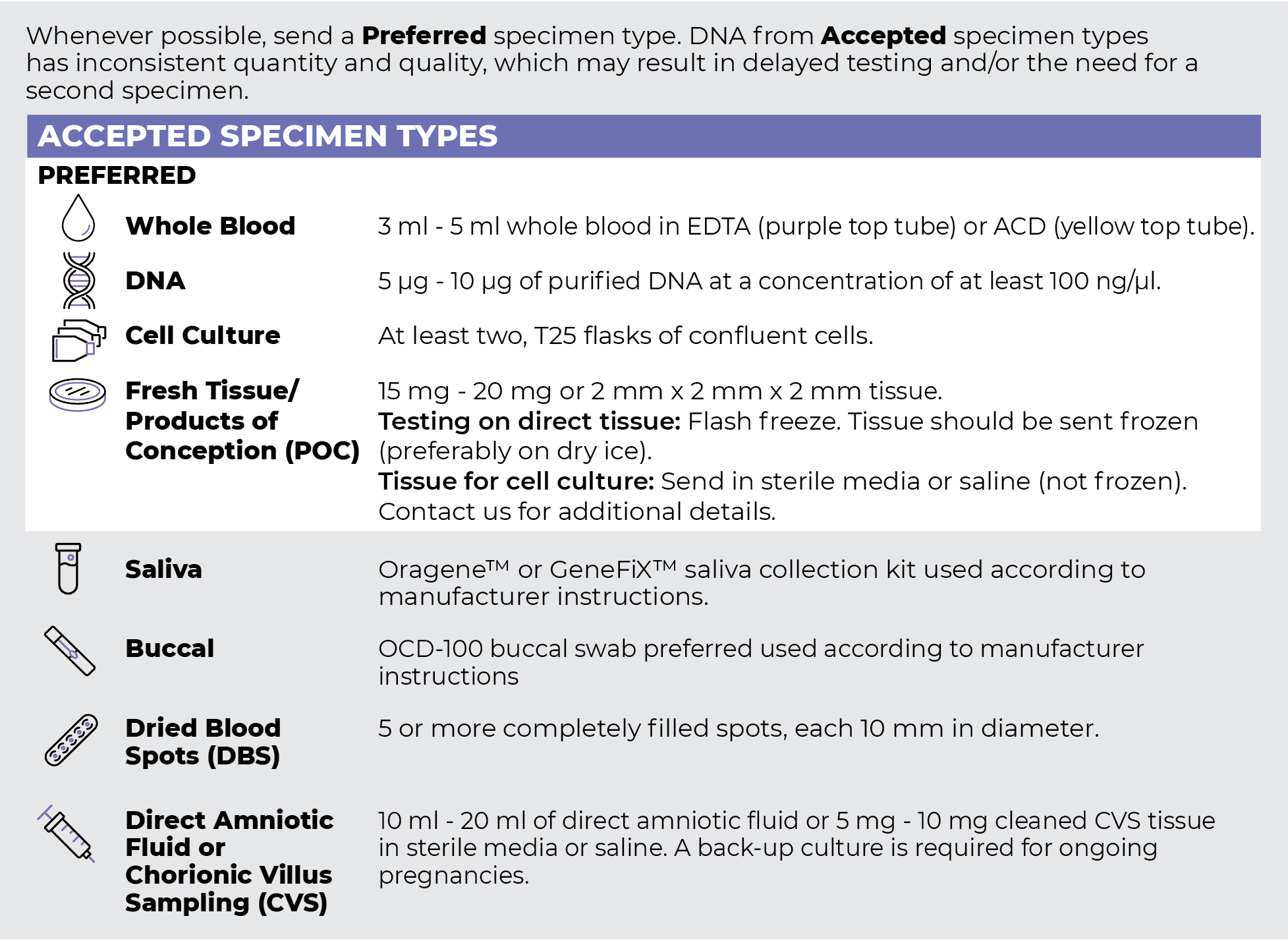
PGnome (Genome) Sequencing Panel

ORDER OPTIONS
View Ordering Instructions1) Select Test Type
2) Select Additional Test Options
No Additional Test Options are available for this test.